文章题目:A plasma membrane transporter coordinates phosphate reallocation and grain filling in cereals
磷(Pi) 对植物生长和作物产量至关重要。然而,在籽粒灌浆过程中 Pi 稳态是如何维持的还不清楚。在本研究中,何祖华课题组通过图位克隆鉴定了一类关于水稻籽粒灌浆调控 Pho1 -Pi 的品系:OsPHO1;2。且发现该品系在灌浆过程中 Pi 重新分配中的特殊作用有关。 伯豪生物为何祖华课题组提供了 RNA-seq 服务,发现 PHO1;2 通过影响 AGPase 的活性进而影响水稻灌浆过程中 Pi 重新分配的作用机制。AGPase 对淀粉合成至关重要,而在 Ospho1;2 突变体植株中,AGPase 的过表达缓解了籽粒灌浆缺陷。该机制在玉米中得到了证实, 为在谷物生长过程中以较小的磷肥投入来提高籽粒产量,提供了有效方案。
籽粒灌浆突变体 OsPHO1;2 的表型及图位克隆
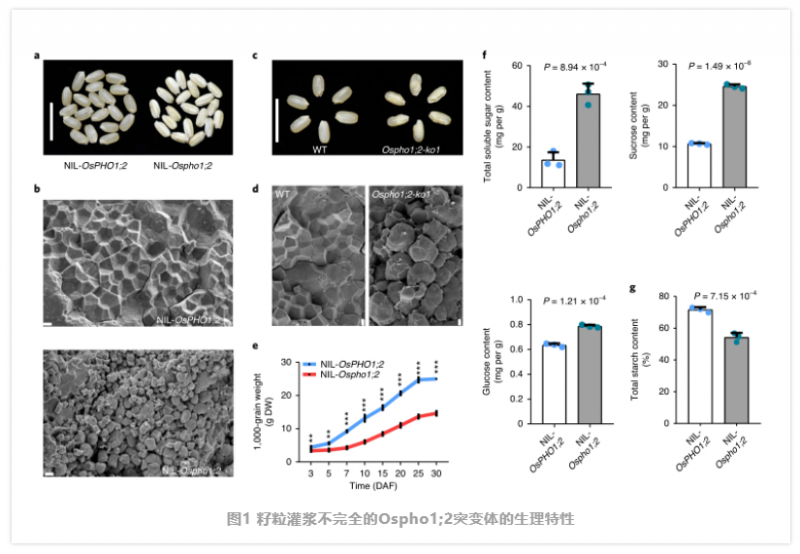
何祖华课题组发现了一个自然发生的籽粒灌浆较差表型的水稻突变体品系。该缺陷表型由一个隐性位点控制,经过基因克隆后分别命名为 NIL-OsPHO1 和 NIL-OsPHO2。NIL-OsPHO2 的籽粒表现出典型的灌浆缺陷,淀粉粒松散、排列不整齐,导致籽粒变薄、皱缩,造成严重的产量损失。除分蘖数减少(图 1) 外,其余突变体植株正常。在整个种子发育过程中,植株的千粒重减少了近一半(图 1e)。与 gif1 突变株不同,NIL-OsPHO1 的淀粉含量更少,可溶性糖更多,对白叶枯病的抗性更强(图 1)。这表明,这两种突变体具有不同的生理表型。在该区域作者鉴定了一个单基因 LOC_Os02g56510,该候选基因编码 PHO1 家族的 Pi 转运蛋白。为了确定 NIL-OsPHO1;2 的原因突变,作者对其 5kb 区域进行了测序,发现在 OsPHO1;2 的第 7 外显子中有 1 -bp 的缺失,导致编码序列的帧移和过早终止。作者还在第 1 外显子和第 3 外显子中发现了两个同义 SNP,在第 12 内含子中发现了一个 SNP。为了确定这一缺陷灌浆表型是否是由 OsPHO1;2 突变引起的,作者在 Nipponbare (NIP) 中构建了 8 个 CRISPR/Cas9 敲除株系,分别为 OsPHO1;2 -ko1 到 OsPHO1;2 -ko8,这些株系均表现出与 NIL-Ospho1;2 相同的灌浆缺陷(图 1)。
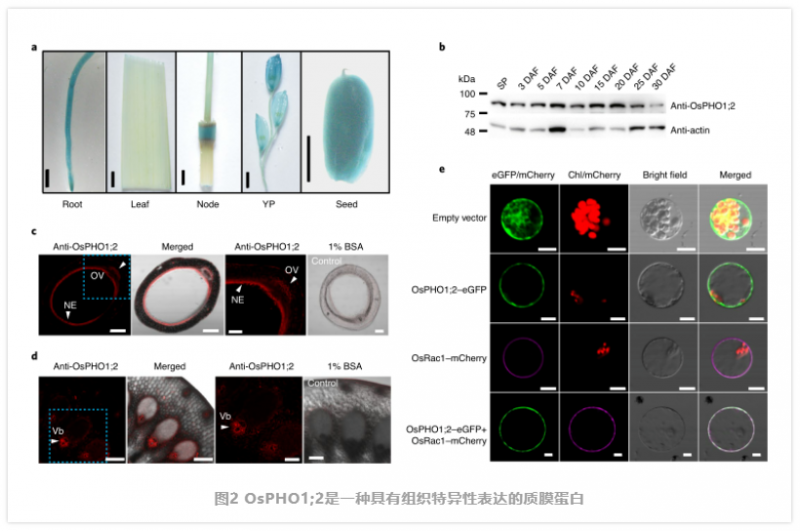
PHO1;2 是一种在种子发育过程中高度表达的质膜蛋白。通过定量反转录 PCR (qRT - PCR) 发现,除了在幼穗、外壳和根中表达外,OsPHO1;2 在发育中的种子中也有高表达。通过对转基因植物中 2 -β- 葡萄糖醛酸酶(GUS) 报告基因的组织学分析,证实了该表达模式(图 2a)。同样,从开花后 0 d(穗)到 30 d( 穗)的整个灌浆阶段,OsPHO1;2 蛋白均在籽粒中富集。为了进一步研究 OsPHO1;2 的细胞特异性,作者在籽粒灌浆早期的种子样本中进行了抗 OsPHO1;2 抗体的免疫荧光试验。有趣的是,在种子的珠心表皮(NE) 和胚珠脉管系统(OV) 中检测到强烈的荧光信号,这些细胞起着向胚乳装载和排出营养物质的作用 m3(图 2c)。在 I 节维管束中也发现了荧光信号(图 2d),这有助于水稻植株中的养分分布。作者进一步验证了 OsPHO1;2 在发育中的种子不同组织中的表达,发现在胚胎、胚乳、果皮和 NE 中都可以检测到 OsPHO1;2 mRNA 和 OsPHO1;2 蛋白。此外,利用含有 pOsPHO1;2- Gus 报告基因的转基因株系进一步验证了 PHO1;2 在种子中的定位。作者将一个瞬时表达的绿色荧光蛋白(GFP) 与 OsPHO1;2 融合进入水稻鞘原生质体和洋葱表皮细胞,发现在这两种情况下,OsPHO1;2 - GFP 主要定位于质膜(图 2)。细胞内偶尔可见微弱的荧光信号,提示部分 OsPHO1;2 蛋白可能定位于细胞内隔间。综上所述,这些结果表明,OsPHO1;2 是一种质膜定位蛋白,在发育中的种子 NE 和维管组织中高表达。
OsPHO1;2 是 Pi 的转运体
由于没有直接证据表明植物 Pho1 型蛋白,如 OsPHO1;2 等是 Pi 的转运体。值得注意的是,虽然 Ospho1;2 幼苗比野生型(WT) 幼苗生长早期,可能由于缺陷胚乳提供的能量较少,它们后来恢复了 WT 的高度。这些生长动态表明,OsPHO1;2 可能不是整个植物生长发育所必需的,但在籽粒灌浆过程中是必不可少的。为了分析 OsPHO1;2 蛋白在籽粒灌浆和 Pi 稳态中的作用机制,作者试图通过三种不同的异源表达系统来确定 OsPHO1;2 蛋白是否确实作为 Pi 转运体。首先,作者发现 OsPHO1;2 补充了酵母突变体 EY917 的生长表型,该突变体缺乏 5 个负责 Pi 摄取的转运体(pho84∆,pho87∆,pho89∆,pho90∆,pho91∆),这证明了 OsPHO1;2 可能具有 Pi 转运活性。主要的外排活性可能介导了木质部的 Pi 负荷,从而解释了先前的遗传分析显示它们在根到地上部 Pi 转移中的作用。为了将 OsPHO1;2 在 Pi 转运中的作用与 Pi 稳态联系起来,作者比较了近距离接触的幼苗之间的 Pi 水平。在缺 Pi 或正常供应 Pi 条件下,突变株根系中 Pi 含量增加,而地上部 Pi 含量降低。因此,OsPHO1;2 突变损害了 Pi 从根到茎的转移,这与之前的结果一致。此外,31P NMR 分析显示,突变体幼苗根细胞中胞质和液泡中 Pi 含量明显较高,排除了 OsPHO1;2 参与胞质 - 液泡划分的可能性(图 3)。接下来,作者研究了成熟期植株地上部组织中 Pi 的分布,发现在节、壳和糙米中,NIL-Ospho1;2 和 Ospho1;2-ko1 植株中 Pi 的含量均高于 WT 植株,但叶片中 Pi 的含量较低。表明 OsPHO1;2 可能参与了 Pi 在种子和叶片之间的重新分配。
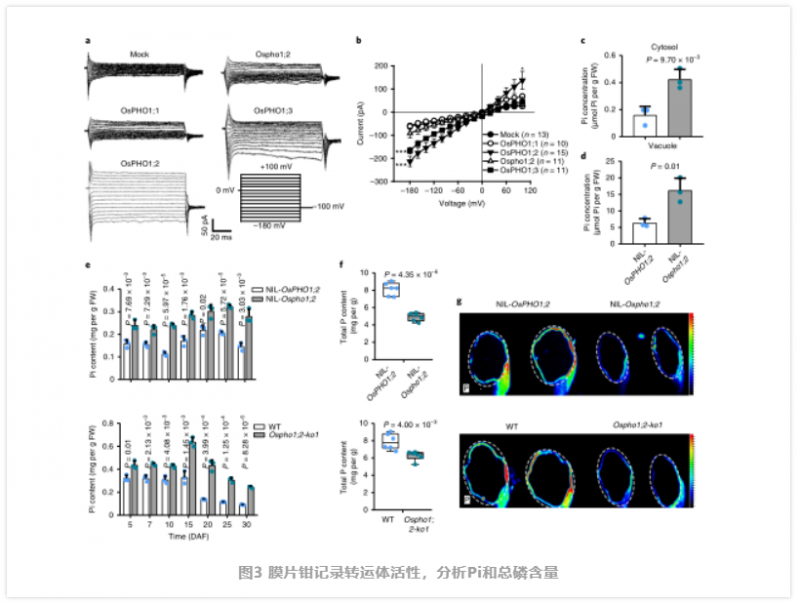
为了支持上述假设,作者进行了测量在整个灌浆过程中发现该突变体的 Pi 含量较高(图 3e)。由于 OsPHO1;2 在发育中的种子 NE 组织中高度表达,我们比较了 NE 周围胚乳和果皮中 Pi 的含量,发现在 Nilosho1;2 种子中,这些组织中 Pi 的含量显著增加。因此,在 Ospho1;2 突变体中,Pi 的再分配受到了干扰。而突变体种子中总磷含量显著降低(图 3f),微 x 射线荧光(μXRF) 成像结果一致(图 3g)。这说明突变株的有机磷合成可能由于种子中母、子组织间 Pi 交换的中断而减少。因此,我们得出结论,OsPHO1;2 作为 Pi 外排转运体,将 Pi 从胚乳细胞中释放出来,其功能的丧失导致 Pi 在种子中过量。过量的 Pi 积累抑制淀粉合成酶。为探讨灌浆期 Pi 积累与灌浆之间的关系,作者对灌浆期 NILs 进行了 RNA-seq 比较分析,鉴定出大量差异表达的基因。作者发现许多参与淀粉和糖代谢的基因被富集。此外,作者通过 qRT-PCR 确定了几个与淀粉合成相关的代表性基因的表达模式,包括编码 AGPase、淀粉合酶、淀粉分支酶和淀粉去分支酶 29(图 4a)。这些基因在 Ospho1;2 突变体中大部分被下调,这与该突变体在淀粉合成方面的缺陷相一致。免疫印迹和同工酶分析显示,突变体种子中参与淀粉合成的几个关键酶的蛋白水平或酶活性也有所下降(图 4)。在这些酶中,AGPase 催化淀粉合成的步骤一,从 G -1- P 和 ATP 中生成 ADP-Glc 和 PPi, AGPase 的功能障碍会导致籽粒灌浆缺陷,这与 Ospho1;2 突变体中发现的缺陷类似,表明 AGPase 在籽粒灌浆过程中起着不可或缺的作用。
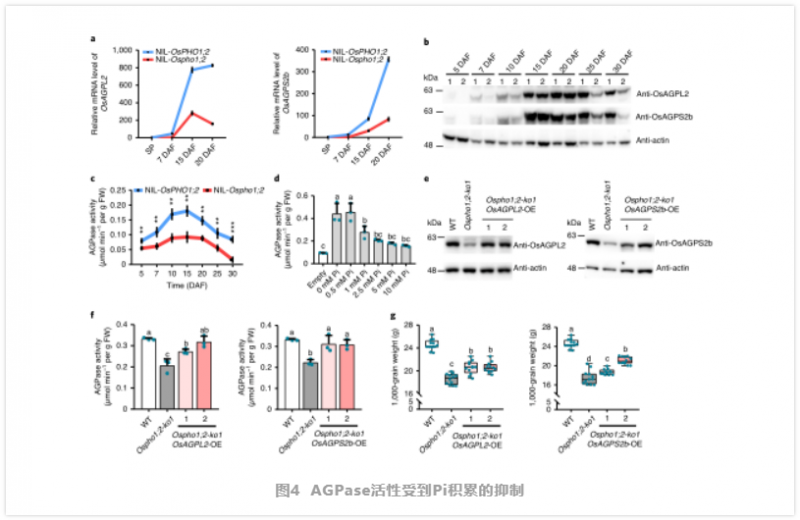
综上所述,作者发现过量的 Pi 水平对 AGPase 的 mRNA 水平和活性都有负面影响,这可能是 Ospho1;2 突变体淀粉合成减少和籽粒灌浆缺陷的基础。为了进一步阐明 AGPase 活性降低与籽粒灌浆缺陷之间的因果关系,作者在 Ospho1;2-ko1 背景下的玉米泛素启动子驱动下,异位过表达了 OsAGPL2 和 OsAGPS2b。选择两个过表达每个转基因的纯合子系进行实验,每个纯合子系的蛋白质水平都恢复到相对于 WT 的水平。作者发现,AGPase 过表达(OE) Ospho1;2-ko1 系的 AGPase 活性低于 WT,但高于 Ospho1;2-ko1 突变体的 AGPase 活性(图 4)。在收获后,我们观察到 AGPase-OE Ospho1;2-ko1 系的灌浆缺陷得到了部分补救,而过表达 OsAGPL2 或 OsAGPS2b 可以显著增加千粒重(图 4)。因此,这些结果进一步支持了 ZmPHO1;2 在籽粒灌浆和 Pi 重新分配中起着重要作用。
伯豪生物服务产品助力国内外临床科研单位发表 SCI 论文 1900 余篇,杂志总影响因子超过 10000 分,其中 Nature、Science、Cell、Lancet Oncol 等高水平论文等高水平论文超过 50 篇。影响因子大于 5 分的文章 659 篇,影响因子大于 10 分的文章 255 篇,影响因子大于 20 分的文章 78 篇,文章领域涉及肿瘤、免疫、神经、发育等多个领域。

更多伯豪生物人工服务:

