
【文章题目】Opposing Wnt signals regulate cervical squamocolumnar homeostasis andemergence of metaplasia
【杂志名称】Nature Cell Biology(IF=17.73)
【发表时间】2021 年 1 月 18 日
【主要技术】10XGenomics
主要结果
鳞状上皮和柱状上皮细胞来源不同
宫颈连接宫腔和阴道,由宫颈外和宫颈内两部分组成。宫颈内壁的柱状上皮与子宫腔相邻,由内口隔开,而宫颈外的鳞状上皮伸向阴道。外口标志着从宫颈外到宫颈内的过渡,在那里柱状上皮与鳞状上皮相遇,形成鳞状柱状上皮交界处(SCJ)(图 1)。
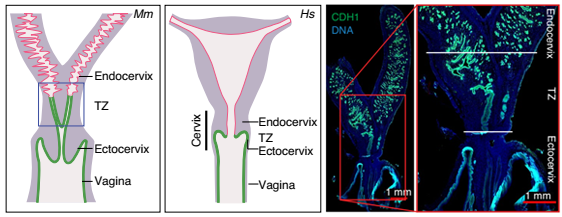
▲图 1 小鼠(Mm)和人(Hs)女性生殖道(FGT)的示意图
作者对健康小鼠的宫颈内膜、宫颈外和过渡区这 3 个组织区域进行 ScRNA-seq 和聚类分析,根据细胞特异性的 marker 基因将亚群分成 7 种类型:基质细胞(Stromal),平滑肌细胞(SMC),周细胞(Pericytes),神经细胞(Neural),免疫细胞(Immune),上皮细胞(Epithelial),内皮细胞(Endothelia)(图 2b,c)。
聚类结果发现宫颈内的细胞类型与过渡区的细胞类型相似,但宫颈外的上皮细胞(Epithelial)、基质细胞(Stromal)、免疫细胞(Immune) 和平滑肌细胞(SMC) 亚群显示出不同的转录谱(图 2a)。
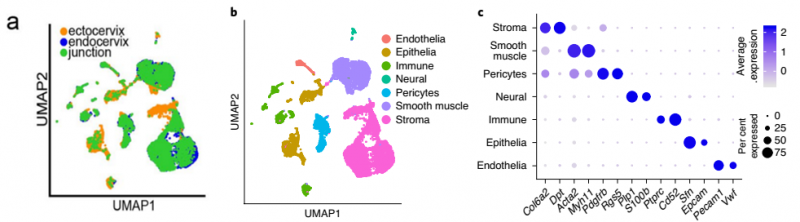
▲图 2 健康小鼠的宫颈内膜、宫颈外和过渡区组织的 scRNA-seq 结果
对上皮细胞群体进行重新聚类,揭示了两种主要的上皮细胞类型:Krt5hi/Krt14hi 鳞状细胞(SQ) 和 Krt8hi/Krt19hi 柱状细胞(Co)。这些进一步被分为六个转录上不同的亚群(SQ1、Sq2A、Sq2B、SQ3、CO1 和 CO2),其中将肌上皮细胞(Me) 归类为 Krt5hi/Krt14hi 亚群,它与 KRT5+ 细胞和肌成纤维细胞有相同的基因表达谱。同时也表达成纤维细胞 marker 即 Col6a2。这些细胞可能是鳞状细胞系的候选前体(图 3)。
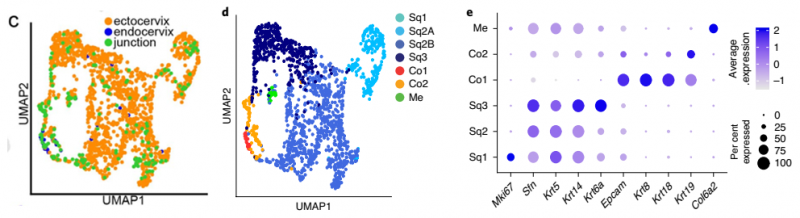
▲图 3 健康小鼠的宫颈内膜、宫颈外和过渡区组织上皮细胞细分结果
SQ1 亚群含丝分裂相关基因,表达 Trp63、Birc5、Mki67、Cks2 和 Hmgb2,表明复层宫颈的基底干细胞。Sq2A 和 Sq2B 亚群含皮肤发育和应激或损伤反应基因,并表达 Trp63、Krt15、Dkk3 和 Notch1。SQ3 亚群含角质形成细胞分化和皮肤发育基因,表达 Fam25c、Gm94、Krt6a 和 Krt10,表明鳞状细胞分化。
CO1 亚群含分支上皮形态发生和脂质分解过程相关基因,表达 Anpep、Cxcl17、Krt8 和 Krt18。CO2 亚群含应激反应、细胞凋亡和对未折叠蛋白的反应基因,并表达 Krt19、Ltf、Muc1 和 Psca(图 3)。
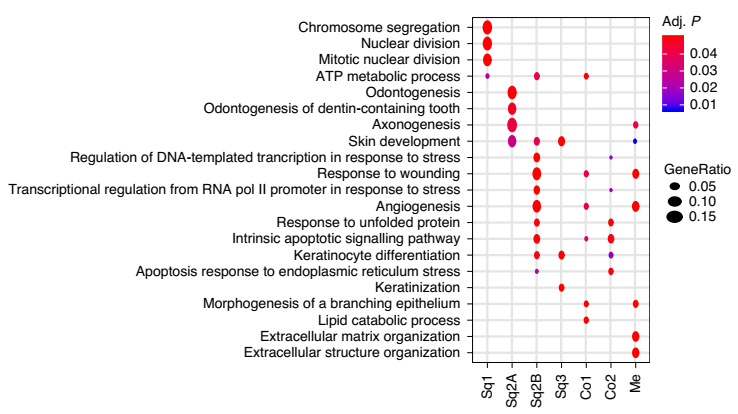
▲图 3 上皮细胞亚群的相关 GO 富集结果
荧光免疫组织化学和 smRNA-ISH 证实 KRT5 和 KRT8 分别标记了整个鳞状分层和柱状上皮细胞。作者通过 Krt5-CreERT2;Rosa26-tdTomato 和 Krt8-CreERT2;Rosa26-tdTomato 小鼠进行谱系追踪,Krt5+ 细胞仅标记复层上皮,而 Krt8+ 细胞仅标记宫颈内上皮。两种上皮在过渡区融合,而 Krt5+ 鳞状细胞似乎取代了覆盖 Krt8+ 的柱状细胞。结果表明这两种上皮细胞类型来自两个不同的谱系(图 4)。
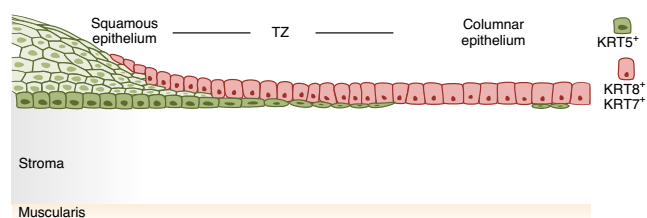
▲图 4 宫颈的鳞状和柱状谱系以及过渡区的示意图
过渡区的上皮边界由不同的基质 Wnt 信号界定
作者通过类器官模型来确认维持这两种上皮细胞的干细胞微环境是不同的。在 Wnt- 的培养基(不含 Wnt3a 和 Rspo1) 中培养的宫颈内膜上皮细胞产生 P63+,Krt5+ 复层的类器官,类似于宫颈外类器官。源自宫颈外的细胞仅产生鳞状类器官,说明与 Wnt 无关。而柱状类器官的形成是依赖于 Wnt 的,与鳞状类器官的形成不同。类器官的 Microarray 分析表明,Wnt 在宫颈内细胞和宫颈外细胞中的转录调控有所不同:Wnt 激动剂在柱状上皮中上调,而 Wnt 拮抗剂 Dkk1 和 KREMEN1 在鳞状上皮中上调(图 5)。
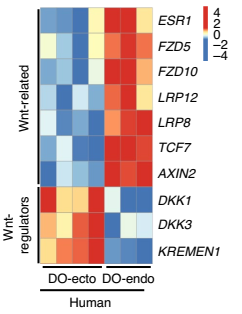
▲图 5 人宫颈外与宫颈内类器官差异调控基因的表达分析
为了深入了解基质如何影响这两种不同的宫颈上皮的动态平衡,作者结合 scRNA-seq 数据的聚类分析揭示了基质细胞群的五个转录上不同的亚群。S1,S2 和 S3 亚群代表宫颈内膜移行区的细胞,通常表达 Hes1,Nfkbia 和 Egr1。S2 亚群高表达 Wnt16 和 Axin2,S3 亚群高表达 Mmp3,Adh1,Ces1d,Cxcl16,Rspo1 和 Rspo3。亚群 S4A 和 S4B 代表宫颈基质细胞;亚群 S4A 显示 Sfrp1 和 Bmp4 的优先表达,亚群 S4B 表达 Dkk2 和 Crabp1。smRNA-ISH 分析也证实了 scRNA-seq 数据(图 6)。
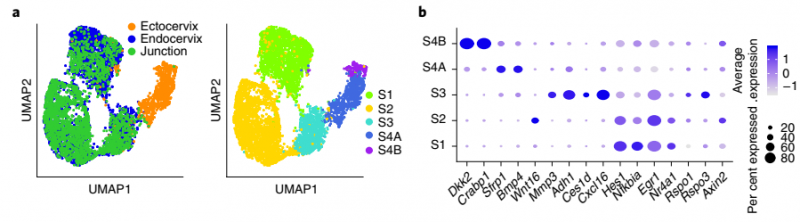
▲图 6 健康小鼠的宫颈内膜、宫颈外和过渡区组织基质细胞的重新聚类结果
为了进一步研究 DKK2 和 DKK3 的作用,作者将宫颈内膜类器官重新接种在含有 WNT3A 和 RSPO1 加 DKK2/DKK3/DKK2 和 DKK3 的培养基中。将 Wnt 缺乏的培养基用作对照。DKK2 的存在导致柱状类器官大小的显着减少,类似于在 Wnt 缺乏的培养基中发现的那样,这证实 DKK2 发挥了抑制作用。当培养基中还含有 DKK3 时,可以避免这种情况。DKK3 要么对 Wnt 信号没有影响,要么作为 Wnt 激动剂发挥作用。用 Wnt 抑制剂 2(IWP2) 阻断 Wnt 信号,抑制宫颈内类器官生长,但不影响宫颈外类器官。这表明鳞状上皮不依赖于细胞自主的 Wnt 信号。
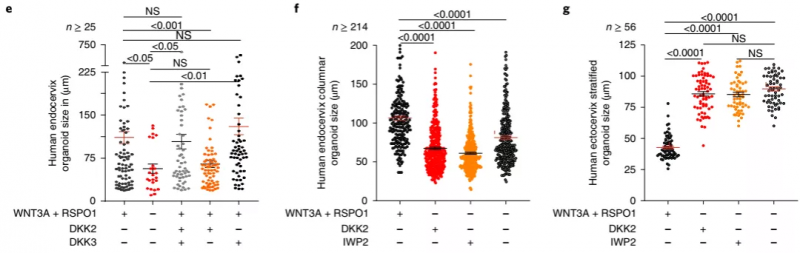
▲图 7 人宫颈内膜柱状类器官和宫颈外复层类器官大小
用 DKK2 或 IWP2 处理后,柱状和鳞状类器官中谱系特异性标记 KRT5,KRT8 和 p63 的表达没有差异。总之,这些数据表明宫颈的上皮由两个不同的干细胞群体维持,它们的命运由相反的 Wnt 信号微环境决定,在过渡区具有明确的开关(图 8)。
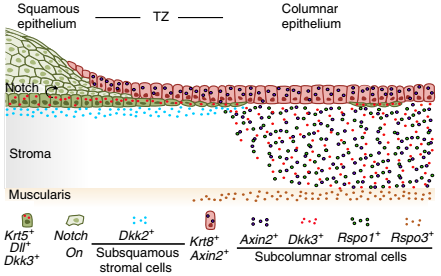
▲图 8 过渡区不同上皮谱系和组织微环境的示意图
WNT 拮抗剂、Notch 和 EGFR 信号通路维持宫颈外干性和分化
基因芯片分析显示,Notch 相关基因在宫颈外鳞状类器官中的表达高于在宫颈内柱状类器官中的表达(图 9)。
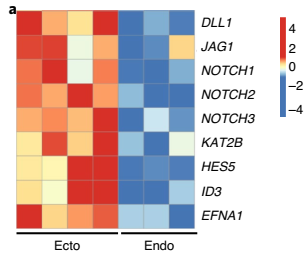
▲图 9 人宫颈外和宫颈内类器官差异基因热图
作者通过对 2D 生长的宫颈外胚层细胞(2D-ECTO)、三天大的早期类器官(EO-ECTO) 和两周大的成熟类器官(DO-ECTO) 进行比较分析来确认分化的机制。结果表明 2D-ECTO 和 EO-ECTO 的基因表达谱具有很高的相似性,但与 DO-ECTO 相比有着明显差异(图 10e)。同样的,2D-ECTO 和 EO-ECTO 与干细胞 marker 的基因集中的表达谱有很高的相似性,但成熟的 DO-ECTO 则不同(图 10f),这说明 2D-ECTO 和 EO-ECTO 的细胞对应于宫颈外干细胞。对宫颈外干细胞相对于分化细胞上调的基因的调查显示,Notch 配体 DLL3 和 MFNG 高表达。而分化的细胞高表达 NOTCH2 和 NOTCH3 受体及其靶点,包括转录因子 HES1 和 γ 分泌酶的核心成分 PSEN1。宫颈外干细胞也表达 Dkk1 及其受体 KREMEN2(图 10g)。因此,使用 γ - 分泌酶抑制剂 DBZ 阻断 Notch 激活会抑制类器官的生长,无法分化和分层。由此表明宫颈干细胞充当 Notch 信号的来源,而分化细胞充当 Notch 信号的接收细胞,其是促进分化和上皮分层的反式激活相互作用。
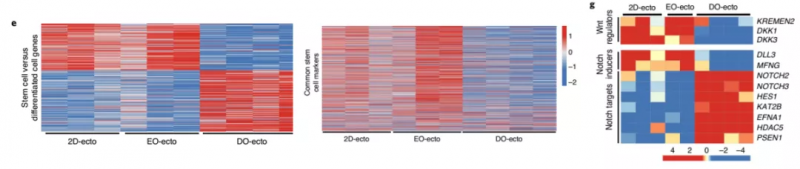
▲图 10 2D-ECTO、EO-ECTO、DO-ECTO 差异基因热图
GSEA 富集显示,受 Notch 下游转录因子和 EGFR-Ras-MAPK 信号转导调控的基因在宫颈外干细胞中高度富集,包括转录因子 AP1、CREB、ETS、NERF、ELK1、E2F、SRF、MYC 和 YY134-37(图 11)。这两条途径共同调节增殖和分化,与表皮生长因子在层状类器官形成中的重要作用一致。相反,属于 RAS 拮抗的 NF1 通路的基因在分化细胞中富集。这些发现表明,Wnt 拮抗剂与 EGFR 和 Notch 诱导通路共同调节宫颈外干性和分化。
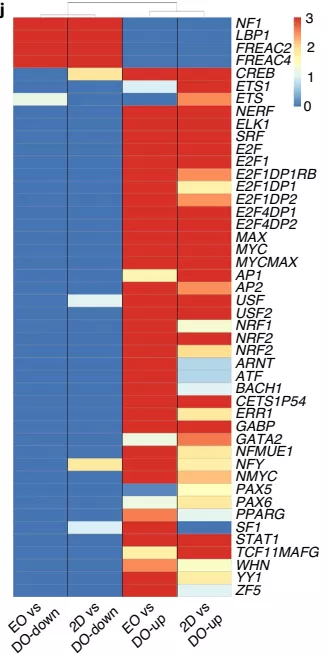
▲图 10 GSEA 富集共有转录因子的顺式调控 motifs 相关基因热图
基质区室的重塑导致 Krt5+ 鳞状化生的出现
为了研究与宫颈外鳞状上皮的干性和分化有关的信号通路和细胞类型在鳞状化生中的作用。作者将人子宫内膜来源的柱状细胞放在 Wnt+ 的培养基中培养时,会产生少量的 Krt5+ 或 p63+ 细胞,同样培养基转移到类器官条件下会产生柱状类器官;然而在 Wnt- 的培养基中培养时,出现对 Krt5+ 和 p63+ 细胞的显著富集的情况,在转移到类器官条件下,即使在 Wnt3a 和(或)Rspo1 存在的情况下,这些细胞也只产生鳞状类器官。相比之下,之初 Wnt+ 或 Wnt- 的培养基中生长的人宫颈外细胞,在转移到 Wnt- 的培养基后只产生鳞状类器官。这表明子宫内膜中存在鳞状干细胞,在适当的微环境下可以产生 Krt5+p63+ 细胞。
作者通过诱导 Krt5-CreERT2;Rosa26-tdTomato 和 Krt8-CreERT2;Rosa26-tdTomato 小鼠的谱系追踪证实了柱状上皮和复层宫颈上皮来自两个不同的谱系,每一个谱系都对特定的微环境做出反应。改变 Wnt 信号不会诱导柱状细胞向层状细胞的转分化,反之亦然。
为了分析体内的化生发展,研究人员利用了缺乏维生素 A 的饮食喂养的小鼠的模型,在该模型中,鳞状化生的异常病灶出现在子宫内膜。为了确定上皮和基质的变化,他们对从宫颈内、宫颈外、过渡区和宫颈内化生(即化生灶)分离的细胞进行了 scRNA-seq 和聚类分析(图 11)。
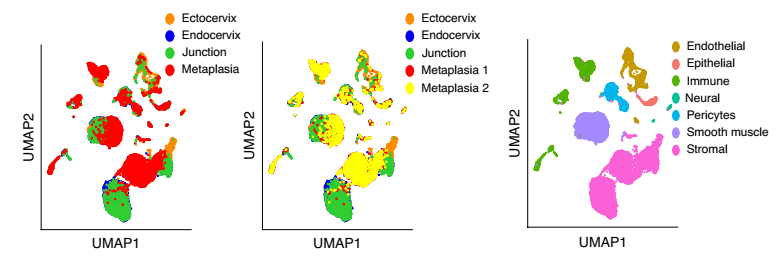
图 11 健康小鼠宫颈外,宫颈内,过渡区和 VA 缺乏引起的鳞状化生的 scRNA-seq
上皮群体被细分成 Sq,Co 和 Me 亚群,根据每个上皮亚群的表达谱生成的系统发育树显示:鳞状细胞亚群(SQ1、Sq2A、Sq2B 和 SQ3) 和 Me 细胞(Me) 在转录上相似,但与柱状亚群(CO1 和 CO2) 不同。Sankey 图是用来分析每个组织区域的上皮细胞对不同亚群的转录贡献,结果表明与健康的宫颈内膜上皮相比,宫颈内膜化生表现出与鳞状上皮和肌上皮型相似的表达谱的扩增(图 12)。
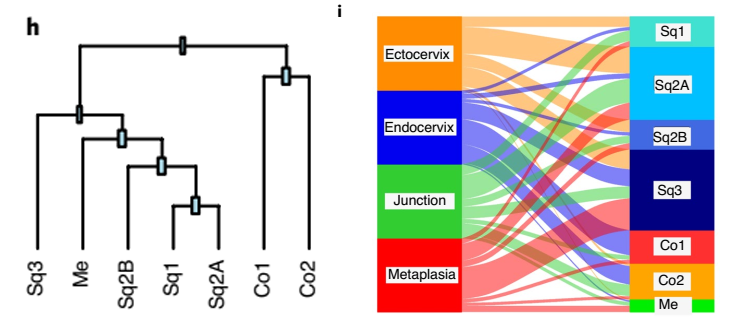
▲图 12 上皮亚群的表达谱生成的系统发育树和 Sankey 图
作者通过对宫颈内、宫颈外、移行区和宫颈内化生的基质细胞进行重新聚类来描述基质细胞亚群的变化(图 13a)。宫颈上皮内化生的基质部分与健康的宫颈内、宫颈外和移行区的基质细胞有明显的差异(图 13b)。,Sankey 图同样表明了宫颈内化生(S2B、S2C 和 S2D) 中出现了与健康宫颈以及外宫颈不同的基质细胞亚群(图 13c)。作者还观察到 Dkk2 在宫颈内化生的一个亚群中表达增加,这主要对应于健康小鼠的宫颈外 S4B。这些数据表明,在化生发育过程中,基质发生了广泛的改建(图 13e)。
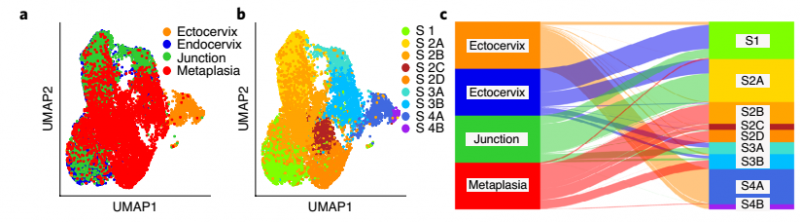
▲图 13 健康小鼠宫颈外,宫颈内,过渡区和 VA 缺乏引起的鳞状化生基质细胞 t -SNE 和 Sankey 图
宫颈癌 SCC 和 ADC 的基因表达模式与鳞状和柱状器官特征相关
成体干细胞易转化,通常构成各种癌症的起源细胞。到目前为止,尚不清楚 ADC 和 SCC 起源于哪种细胞类型。作者根据鳞状和柱状宫颈类器官的基因表达特征来确定宫颈癌起源的可能细胞。TCGA 中的基因表达数据将宫颈癌分为 ADC 和高角蛋白 SCC 组和低角蛋白 SCC 组,其中低角蛋白 SCC 组显示鳞状上皮特异性标记物的较低表达。作者根据宫颈外鳞状上皮和宫颈内柱状上皮之间差异表达的基因,将癌症样本分为鳞状上皮样和柱状上皮样。结果发现分类为柱状样或鳞状样的癌症与分类为 ADC 或 SCC 的组织学分类之间存在一致性;这从与 ADC 或角蛋白高和角蛋白低组的 TCGA 簇的比较中也是明显的。
其中一组肿瘤含量低,与宫颈外或宫颈内征象无明显相似性的病例被分类为未确定病例。然而,在 62 例未确定的病例中,有 59 例落在 TCGA 定义的角蛋白低或高角蛋白的鳞状细胞癌丛中。值得注意的是被归类为柱状的癌症样本主要是 KRT5low、KRT7high 和 TP63low,而被归类为鳞状和未确定的癌症样本主要是 KRT5high 和 TP63high,并混有的 KRT7。这表明未确定的组可能由宫颈内或从宫颈内长出的 SCC 组成,导致样本中存在污染的 KRT7+ 宫颈柱状细胞。
研究表明位于过渡区的 SCJ 细胞表达 KRT7、CD63、GDA、AGR2 和 MMP7,这些细胞是 SCC 和 ADC19 的前体细胞。因此作者研究了类器官和 TCGA 宫颈癌样本中的基因表达水平,检测了 SCJ 标记物 Krt5。这些标记物在宫颈内类器官和 ADC 中的表达明显高于在宫颈外类器官和鳞癌中的表达,而 Krt5 的表达趋势则相反。人和小鼠宫颈组织切片的免疫组织化学和 smRNA-ISH 证实 Krt5 在整个鳞状上皮中均有表达。KRT7 在整个柱状上皮中表达,在较小程度上在鳞状上皮中表达,而不是仅限于过渡区。宫颈外类器官对鳞状上皮标记 Krt5 和 CTSA 呈阳性反应,而宫颈内类器官对柱状上皮标记 KRT7、AGR2、GDA 和 Kr18 呈阳性反应。这意味着已报道的 SCJ 细胞与宫颈柱状细胞谱系没有区别,也不是 SCC 的起源细胞。
与角蛋白高和角蛋白低的鳞状细胞癌均起源于鳞状细胞系的观点一致,该研究还揭示了高角蛋白和低角蛋白的鳞状细胞癌和 ADC 组之间的差异表达基因。作者在柱状癌和鳞状癌(包括 MUC5B、KRT5 和 CSTA) 中观察到双模式基因表达,它们将 ADC 与 SCC 区分开来。SCJ 的标记物 KRT7、AGR2 和 GDA 只标记 ADC,而不标记 SCC。综上所述,大多数宫颈癌可以根据分子特征分为两组:SCC 可能起源于宫颈外的 Krt5+ 鳞状谱系,而 ADC 可能起源于宫颈内的 KRT7+KRT8+ 柱状谱系(图 14)。
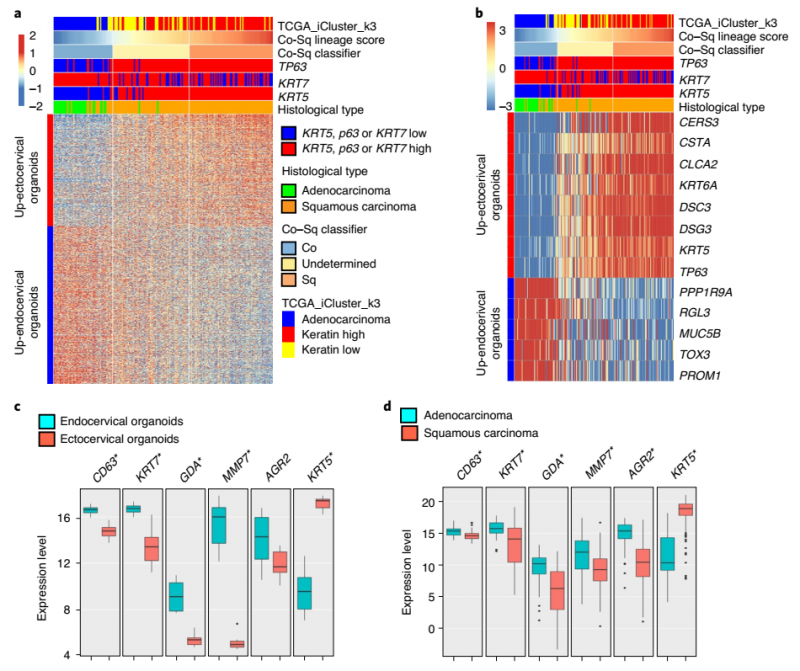
▲图 14 宫颈癌 SCC 和 ADC 的转录特征与鳞状和柱状上皮谱系相关
总结
综上所述,作者的数据表明宫颈鳞状上皮和柱状上皮起源于两种不同的谱系特异性干细胞类型,并确定了这些谱系的亚群。此外,宫颈内外有不同的基质亚群,构成了维持过渡区的微环境梯度。通过信号梯度调节干细胞的维持、分化和组织构型是发育过程中的普遍原则,其中 Wnt 信号是不可或缺的。在基质中存在 Wnt 梯度,它选择性地驱动柱状谱系,同时对同一组织中存在的鳞状谱系特异性干细胞施加静止。在鳞状化生中,宫颈内基质经历了广泛的改建。作者还观察到 Wnt 抑制剂 DKK2 在基质亚群中上调。这种向 Wnt 抑制微环境的转变可能激活静止的鳞状上皮干细胞,这些干细胞侵入移行区的柱状上皮或作为子宫内膜内的化生灶(图 15)。从分子和细胞的角度来看,目前对维持宫颈上皮连接的机制的阐明提供了一个重要的概念进展。这表明,这些部位的动态平衡并不是通过一种上皮细胞类型向另一种上皮细胞类型的转分化来维持的。取而代之的是,成体组织类似于不同谱系特异性干细胞群体的整合,由微环境对外部或内部信号做出反应而激活。粘膜过渡区动态平衡的这一概念与对粘膜干细胞鉴定的其他观察结果非常吻合,并为将来在其他具有高度临床意义的组织中研究过渡区动态平衡提供了基础。
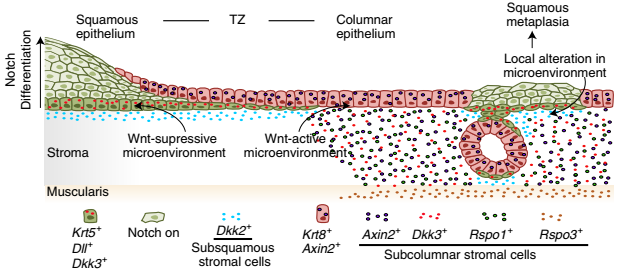
▲图 15 移行区和鳞状化生期间的两种上皮谱系和 Wnt-Notch 微环境
更多伯豪生物人工服务:

